
Séance 1: Anatomie de la tige (structure primaire)
La structure primaire de la tige, formée par le méristème primaire apical ou méristème primaire caulinaire, se divise en plusieurs catégories de tissus essentiels au développement et au fonctionnement des organes végétaux.
Les catégories de tissus des organes végétaux
1. Tissus de revêtement (ou dermes) : Constitués d’une couche unique de cellules, ces tissus recouvrent la surface externe des jeunes plantes, assurant protection et facilitant les échanges avec l’environnement.
2. Tissus fondamentaux : Jouant un rôle prédominant dans les fonctions métaboliques, ces tissus se distinguent par leur couleur jaune sur les schémas et se subdivisent en plusieurs types de cellules :
Parenchyme :
Le parenchyme est un type de tissu végétal composé de cellules généralement larges et volumineuses, qui se caractérisent par leur adaptabilité à remplir diverses fonctions essentielles au sein de la plante. Ces cellules se distinguent par leurs parois fines et souvent arrondies, et sont séparées les unes des autres par des espaces intercellulaires appelés méats.
Ce tissu peut être identifié et classifié de différentes manières, selon sa localisation, son association à d’autres types de tissus, ou encore sa fonction spécifique au sein de l’organisme végétal. Par exemple, on peut parler de parenchyme cortical (situé dans la couche externe du tissu) ou médullaire (situé dans la partie centrale). Lorsqu’il est associé à d’autres types de tissus, on peut le désigner comme parenchyme libérien (lié au liber) ou ligneux (associé au bois). En fonction de sa fonction, le parenchyme peut être qualifié d’accumulateur (stockage de réserves), d’aérifère (transport de l’air) ou de chlorophyllien (participation à la photosynthèse).
Il est important de noter que le parenchyme d’un organe végétal âgé peut subir une transformation, ses cellules lignifiant leurs parois pour devenir un tissu de soutien plus rigide, connu sous le nom de parenchyme lignifié. Cette capacité d’adaptation et de transformation reflète la complexité et la diversité des rôles du parenchyme au sein des plantes, soulignant son importance fondamentale dans leur biologie et leur écologie.
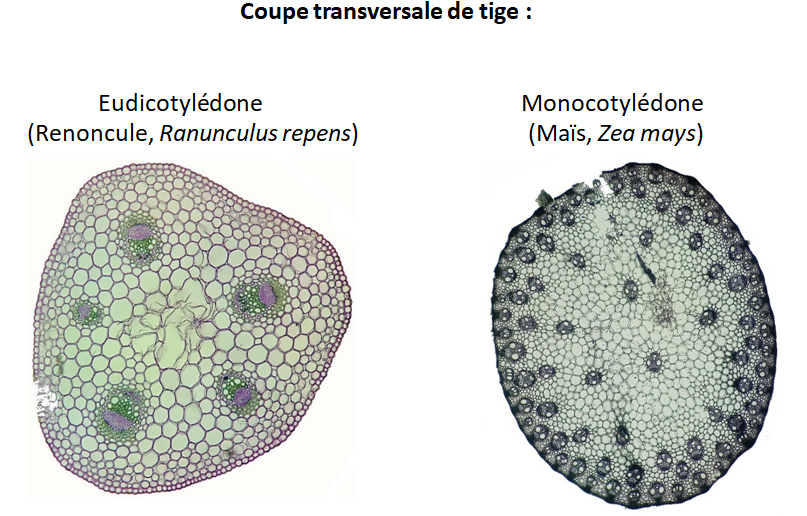
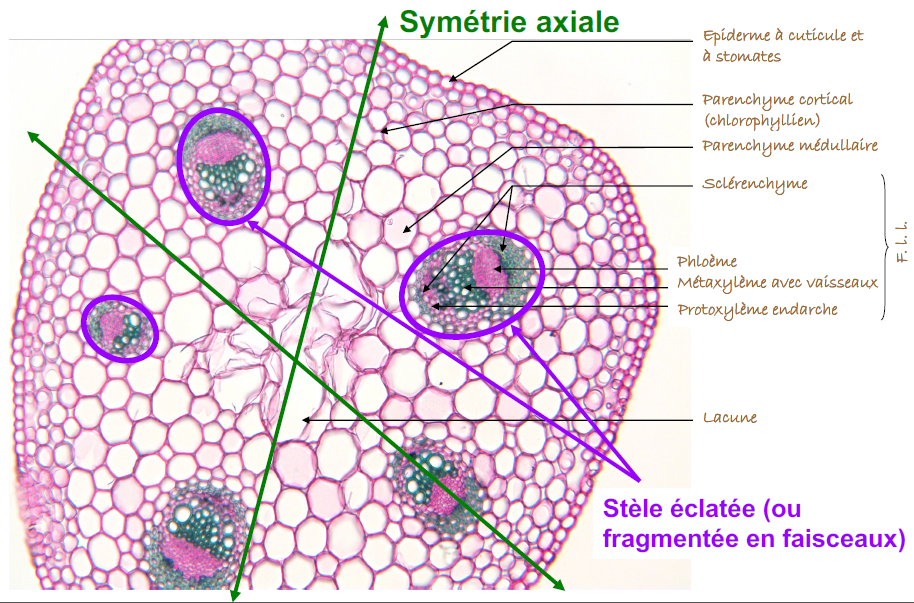
La tige des eudicotylédones : renoncule
Collenchyme :
Le collenchyme est un tissu primaire de soutien vivant et souple, fréquemment retrouvé dans les tiges jeunes et les feuilles, caractérisé par des ponctuations dans les parois cellulaires qui facilitent la circulation des substances entre les cellules. Ce tissu se distingue par l’épaississement variable de ses parois, ce qui donne lieu à différents types de collenchymes en fonction de la localisation de cet épaississement : angulaire lorsque l’épaississement se situe aux angles des cellules, rond si l’épaississement est uniformément réparti, ou tangentiel lorsque l’épaississement se trouve sur les faces parallèles aux surfaces de l’organe. La forme des cellules peut également varier, donnant naissance à des fibres de collenchyme spécifiques. Ces caractéristiques permettent au collenchyme d’offrir un soutien flexible aux parties en croissance de la plante, contribuant ainsi à sa structure et à sa résistance.
Sclérenchyme :
Le sclérenchyme est un tissu de soutien primaire dans les plantes, caractérisé par sa rigidité, sa dureté, et son aspect coriace, résultant de cellules mortes à maturité. Ces cellules, souvent polygonales et étroitement jointives, présentent des parois épaisses ponctuées qui facilitent une circulation limitée. La particularité de ce tissu réside dans son processus de développement : la paroi primaire est initialement renforcée par une paroi secondaire dense en cellulose à l’intérieur de la cellule. Par la suite, la lamelle moyenne, la paroi primaire, et la paroi secondaire s’imbibent de lignine, entraînant la mort de la cellule. Les cellules sclérifiées, typiquement regroupées en petits amas, se manifestent souvent sous forme de fibres extrêmement allongées. Ce processus d’imprégnation de lignine et la mort subséquente des cellules dotent le sclérenchyme de ses propriétés de soutien exceptionnelles, cruciales pour la structure et la résistance des tissus végétaux.
3. Tissus conducteurs : Indispensables pour le transport de l’eau, des nutriments et autres substances, ils diffèrent dans leur organisation selon les organes. Ce groupe inclut :
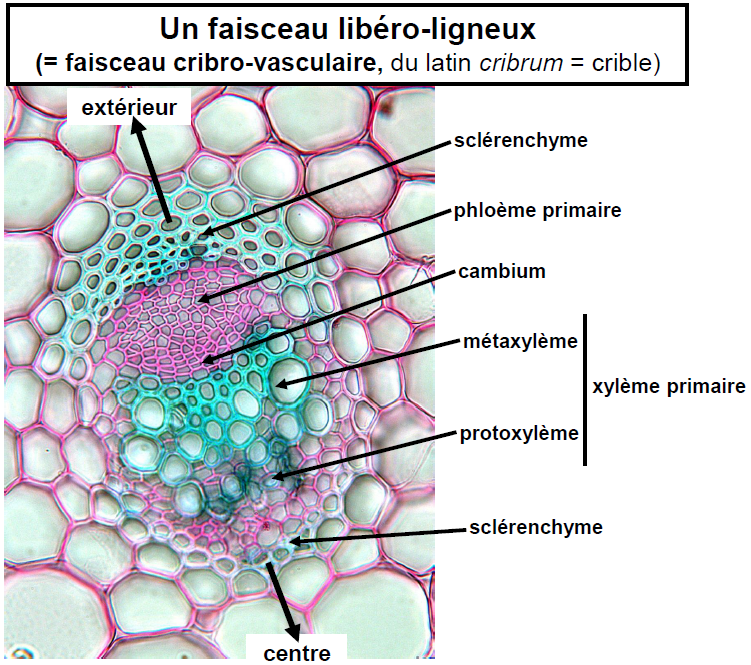
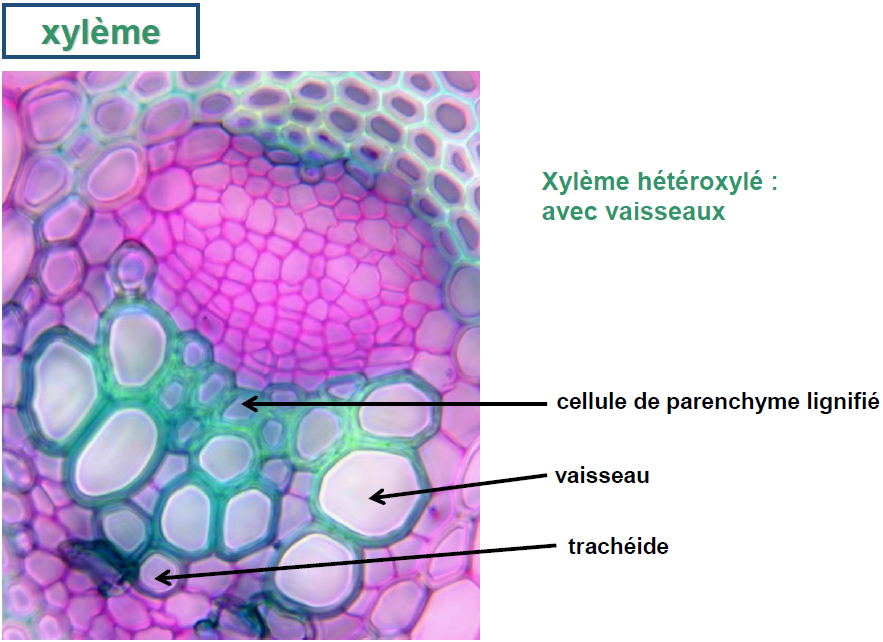
Xylème :
Le xylème, également connu sous le nom de bois, est un tissu végétal essentiel qui remplit plusieurs fonctions importantes. Il est principalement responsable de la conduction de la sève brute, riche en sels minéraux et autres nutriments essentiels, depuis les racines vers le haut de la plante, processus connu sous le nom de mouvement ascendant. Au-delà de son rôle de transport, le xylème sert également de tissu de soutien et de réserve, notamment dans le cas du bois secondaire.
Ce tissu se caractérise par sa composition hétérogène, incluant un mélange de cellules mortes et vivantes, certaines étant lignifiées pour renforcer et soutenir la structure de la plante. Les principaux types de cellules qui le composent sont :
- Les éléments de vaisseaux, qui sont des cellules mortes permettant une conduction verticale rapide de la sève grâce à leurs perforations terminales. Ils sont reconnaissables à leur large diamètre et à leur paroi ponctuée et lignifiée. Ces éléments sont typiquement superposés pour former un vaisseau, une caractéristique exclusive des angiospermes.
- Les trachéides, également lignifiées, assurent une conduction verticale mais plus lente que celle des éléments de vaisseaux, via des ponctuations latérales et terminales. Elles sont particulièrement présentes chez les ptéridophytes et les gymnospermes.
- Les cellules de parenchyme, qui peuvent être cellulosiques ou lignifiées, jouent un rôle physiologique et sont parfois organisées en rayons pour faciliter les échanges et le stockage de réserves.
- Les fibres, longues cellules lignifiées avec un espace interne réduit, contribuent au soutien structurel de la plante.
Le xylème peut être classé en fonction de son origine et de sa structure. Chez les dicotylédones angiospermes, le bois est dit “hétéroxylé” en raison de la présence de vaisseaux, tandis que chez les gymnospermes, il est “homoxylé”, c’est-à-dire sans vaisseaux.
Le développement du xylème se fait en trois phases distinctes :
- Le protoxylème, premier xylème à se différencier dans l’organe en croissance, est identifiable même dans les parties plus âgées de la plante. Ses cellules conductrices, principalement des trachéides avec de la lignine discontinue, permettent un étirement grâce à des configurations annelées, spiralées, réticulées, ou striées, avec une présence notable de parenchyme vasculaire.
- Le métaxylème primaire se développe après le protoxylème, une fois la croissance en longueur de l’organe achevée, pouvant être homoxylé ou hétéroxylé, similaire au bois secondaire mais sans organisation radiale.
- Le bois secondaire ou xylème secondaire se forme à partir du cambium et contribue majoritairement à l’épaisseur des tiges ou des racines des arbres. Il se distingue par l’arrangement radial de ses cellules, incluant les rayons vasculaires, et peut être homoxylé ou hétéroxylé.
Cette structure complexe du xylème reflète son rôle crucial dans la nutrition, le soutien, et la croissance des plantes, soulignant l’ingéniosité de leur adaptation au milieu terrestre.
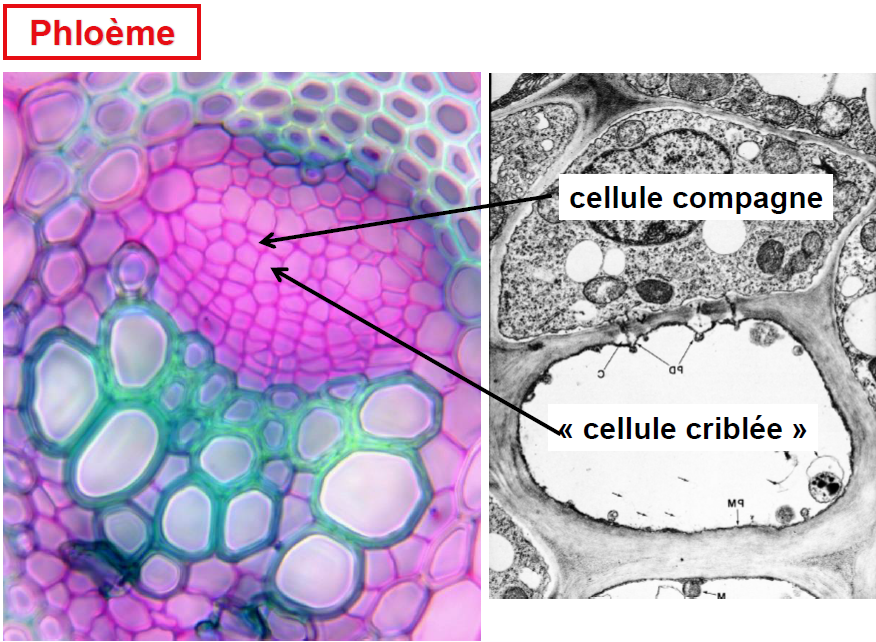
Phloème :
Le phloème, également connu sous le nom de liber, est un tissu conducteur essentiel dans les plantes, responsable du transport de la sève élaborée, riche en sucres, vers le bas de la plante depuis les feuilles où la photosynthèse a lieu. Les caractéristiques distinctives de ce tissu incluent la petite taille de ses cellules, parmi les plus réduites en diamètre au sein d’un organe, et leurs contours polygonaux qui s’assemblent étroitement sans laisser d’espaces intercellulaires (méats).
Le phloème est un tissu hétérogène composé de plusieurs types de cellules :
- Les tubes criblés : Principaux conducteurs de sève élaborée, caractérisés par une conduction rapide grâce à des plages de cribles situées aux extrémités des cellules.
- Les cellules compagnes : Plus petites et associées aux tubes criblés, elles jouent un rôle crucial dans le fonctionnement de ces derniers. Elles ne sont pas présentes chez les Gymnospermes.
- Les cellules de parenchyme libérien : Participent aux fonctions de stockage et de transport au sein du phloème.
- Les cellules de soutien : Telles que les fibres, qui peuvent être cellulosiques ou lignifiées, ajoutant une structure de soutien au tissu.
Le phloème se différencie également selon son stade de développement et son origine :
- Le protophloème : Première forme de phloème à se développer dans les organes jeunes, souvent déformé et non fonctionnel dans les parties plus âgées de la plante, ce qui le rend difficile à reconnaître.
- Le métaphloème primaire : Se différencie après le protophloème, une fois que l’organe a achevé sa croissance en longueur.
- Le phloème (ou liber) secondaire : Se forme à partir d’un cambium secondaire, caractérisé par une organisation cellulaire en files radiales. Il est traversé par des rayons de parenchyme qui facilitent les échanges entre le xylème et le phloème. Ce type de phloème est absent chez les Monocotylédones angiospermes.
Ce système complexe permet au phloème de remplir efficacement sa fonction de distribution des nutriments essentiels à travers la plante, soutenant ainsi son développement et sa croissance.
Cambium :
Le cambium, également désigné sous le terme d’assise génératrice libéro-ligneuse, représente une zone de croissance secondaire essentielle chez les plantes, principalement responsable de la production du bois (xylème) secondaire et du phloème (liber) secondaire. Cette couche génératrice est caractéristique des Angiospermes Dicotylédones et des Gymnospermes, où elle apparaît initialement entre le bois et le phloème primaires, avant de potentiellement s’étendre autour de l’organe.
Le cambium est composé de cellules tabulaires, c’est-à-dire de cellules rectangulaires très allongées et particulièrement fines, qui s’organisent en courtes files radiaires. Ces cellules se distinguent par leurs parois extrêmement fines, permettant une division cellulaire rapide et efficace. Cette structure permet au cambium d’assurer sa fonction de tissu de croissance, en générant de nouvelles cellules qui contribueront à l’épaississement de la plante par l’ajout de couches successives de xylème à l’intérieur et de phloème à l’extérieur.
Le rôle du cambium dans la croissance secondaire est crucial pour le développement structurel des plantes, leur permettant d’accroître leur diamètre annuellement et de renforcer leur capacité de soutien et de transport des nutriments.
Epiderme :
L’épiderme représente le tissu primaire de revêtement des tiges et des feuilles chez les plantes, constituant une couche protectrice vivante. Il est formé d’une assise de cellules aux parois principalement cellulosiques, et, dans certains cas, lignifiées. Ces cellules se situent à la surface externe de la plante et remplissent diverses fonctions essentielles.
L’épiderme se compose principalement de deux types de cellules :
- Les cellules de revêtement : Elles forment la majorité de l’épiderme et servent à protéger la plante contre les agressions extérieures et la perte d’eau.
- Les cellules stomatiques : Cruciales pour la régulation des échanges gazeux et la transpiration, elles peuvent s’ouvrir et se fermer pour contrôler le passage de l’air et de l’eau.
En plus de ces cellules principales, l’épiderme peut comporter des structures spécifiques telles que des poils, qui peuvent être sécréteurs dans certains cas, contribuant ainsi à des fonctions variées comme la protection contre les herbivores et les maladies, ou l’absorption d’eau et de nutriments.
Une caractéristique distinctive de l’épiderme est la présence d’une cuticule sur sa surface externe. Cette couche est composée de cutine et contient des sécrétions lipidiques, incluant des cires imperméables, formant ainsi une barrière efficace contre l’évaporation excessive de l’eau. Cependant, avec l’augmentation du diamètre des tiges due à la croissance secondaire, l’épiderme est souvent remplacé par du tissu périphérique plus résistant, comme le périderme, entraînant la disparition de la cuticule dans ces zones en expansion.
Anatomie comparée des tiges :
- Eudicotylédones (ex. : Renoncule Rampante, Ranunculus repens) présentent une structure cellulaire où la paroi primaire est cellulosique et souple, et la paroi secondaire, plus épaisse, peut être riche en cellulose, lignine, cutine ou subérine. La coloration avec le rouge Congo pour la cellulose et la phloroglucine chlorhydrique pour la lignine est recommandée pour examiner ces structures.
- Monocotylédones (ex. : Maïs, Zea mays) disposent d’un épiderme unique à parois cellulosiques, avec une cuticule cireuse et des tissus de soutien tels que le sclérenchyme pour le support mécanique.
Distinction clé : Les Eudicotylédones montrent un arrangement vasculaire circulaire, tandis que les Monocotylédones présentent des faisceaux vasculaires dispersés, reflétant une complexité d’organisation différente.
4. Classification des tissus végétaux : Les tissus végétaux se classent en deux grandes catégories, selon leur composition cellulaire :
- Tissus simples : Formés d’un seul type de cellules, ils accomplissent des fonctions spécifiques au sein de l’organisme végétal. Le parenchyme, le collenchyme et le sclérenchyme sont des exemples de tissus simples, chacun jouant un rôle distinct dans la plante, que ce soit pour le stockage, le support flexible ou le soutien mécanique durable.
- Tissus complexes : Composés de plusieurs types de cellules différentes, ils assument des fonctions plus élaborées liées au transport et à la distribution de l’eau, des nutriments et des sucres dans la plante. Les tissus conducteurs, tels que le xylème et le phloème, sont des exemples de tissus complexes, indispensables à la survie et au développement de la plante.
Coloration
Après la coloration pour l’observation microscopique, les couleurs des tissus xylème et phloème peuvent varier en fonction des colorants utilisés. Voici des détails sur les couleurs typiquement observées pour chaque tissu après l’application de certains colorants courants :
- Coloration au Safranine-O et Vert de Fast :
- Xylème : Le safranine-O est souvent utilisé pour colorer le xylème, où il prend une couleur rouge. Ce colorant a une affinité pour les parois cellulaires lignifiées, ce qui est caractéristique des cellules du xylème, particulièrement des trachéides et des vaisseaux.
- Phloème : Le vert de fast, utilisé en combinaison avec le safranine-O, colore le phloème en vert. Cela permet de distinguer facilement le phloème du xylème sous le microscope, le vert de fast ayant une affinité pour les parois cellulaires moins lignifiées et plus riches en cellulose, comme celles des cellules du phloème.
- Coloration au Bleu de Toluidine :
- Xylème et Phloème : Le bleu de toluidine peut colorer à la fois le xylème et le phloème, mais il permet de distinguer les structures à l’intérieur de ces tissus en fonction de leur contenu en acides nucléiques et en polysaccharides. Bien que le bleu de toluidine ne différencie pas spécifiquement le xylème du phloème par des couleurs distinctes, il peut révéler des nuances de bleu selon la densité et le type de composants cellulaires, aidant à identifier les différentes structures cellulaires.
- Autres colorants :
- Certains protocoles peuvent utiliser d’autres colorants spécifiques qui se lient préférentiellement aux composants cellulaires du xylème ou du phloème, produisant des couleurs variées selon le colorant et les caractéristiques du tissu. Par exemple, l’utilisation de la phloroglucine-HCl pour détecter la lignine dans le xylème produit une coloration rouge en présence d’acide chlorhydrique.
********************
Le carmino-vert colore la cellulose en rose et la lignine en vert.
Le carmino-vert colore le sclérenchyme en vert.
Le carmino-vert colore le phloème en rose.
La phorogulcine chlorydrique colore en rosele xylème et le sclérenchyme.
Le rouge congo colore la cellulose en rose.
Les différences entre les tissus des Eudicotylédones et des Monocotylédones reflètent des adaptations évolutives distinctes à leurs environnements respectifs. Voici les principales distinctions :
- Disposition des faisceaux vasculaires :
- Eudicotylédones : Les faisceaux vasculaires sont arrangés en un cercle à l’intérieur de la tige, avec un xylème tourné vers l’intérieur et un phloème vers l’extérieur. Cet arrangement permet une croissance en épaisseur grâce à l’activité du cambium, un méristème secondaire situé entre le xylème et le phloème.
- Monocotylédones : Les faisceaux vasculaires sont dispersés à travers le parenchyme de la tige sans arrangement circulaire distinct. Les monocotylédones n’ont généralement pas de cambium, ce qui limite leur croissance en épaisseur.
- Présence et activité du cambium :
- Eudicotylédones : Possèdent un cambium bien développé qui participe à la croissance secondaire, permettant aux tiges de s’épaissir et d’accroître leur diamètre au fil du temps.
- Monocotylédones : La plupart n’ont pas de cambium, ou il est très peu actif, résultant en une croissance en épaisseur très limitée ou absente.
- Structure des feuilles et disposition des nervures :
- Eudicotylédones : Les feuilles présentent généralement des nervures ramifiées ou réticulées.
- Monocotylédones : Les feuilles ont des nervures généralement parallèles.
- Racines :
- Eudicotylédones : Développent typiquement un système racinaire pivotant, avec une racine principale qui pénètre profondément dans le sol, d’où partent des racines secondaires.
- Monocotylédones : Ont tendance à développer un système racinaire fasciculé, composé de nombreuses racines de taille similaire qui se propagent dans le sol, offrant une fixation et une absorption plus dispersées.
- Nombre de cotylédons :
- Eudicotylédones : Les graines contiennent deux cotylédons (feuilles embryonnaires).
- Monocotylédones : Les graines contiennent un seul cotylédon.
Ces différences structurales et anatomiques entre Eudicotylédones et Monocotylédones ont des implications sur la façon dont ces plantes absorbent les nutriments, se développent, et interagissent avec leur environnement. Les adaptations uniques de chaque groupe contribuent à leur diversité écologique et à leur répartition dans différents habitats.
**************************
L’épiderme contient une couche(s) de cellules.
Les cellules du parenchyme sont grosses , rondes et non-jointives.
Le sclérenchyme sert à protéger les tissus.
Quel est le nom du tissu conducteur de sève élaborée ? Phloème
Le xylème contient des trachéides et des vaisseaux chez les angiospermes.
Les cellules du phloème sont vivantes.
De quels types de cellules le xylème est-il constitué?: trachéides et éléments de vaisseaux.
La différence entre Protoxylème et xylème, et Protophloème et phloème
Le protoxylème et le protophloème sont les premières formes de xylème et de phloème qui se développent dans les plantes vasculaires, tandis que le xylème et le phloème matures représentent des stades de développement plus avancés de ces tissus.
Protoxylème
- Définition : Le protoxylème est la première forme de xylème qui se développe dans les plantes. Il apparaît dans les parties les plus jeunes de la plante, comme la pointe de croissance.
- Fonction : Sa principale fonction est le transport de l’eau et des minéraux depuis les racines vers les parties aériennes de la plante. Cependant, étant donné que le protoxylème se trouve dans les régions en croissance, ses éléments sont souvent plus petits et ont des parois minces, ce qui les rend susceptibles de s’effondrer sous la pression.
- Caractéristiques : Le protoxylème est caractérisé par des éléments de vaisseau avec des parois minces et des trachéides, qui peuvent être détruits au fur et à mesure que la plante grandit et que la tige s’épaissit.
Xylème
- Définition : Le xylème est un tissu vasculaire mature dans les plantes, responsable du transport de l’eau et des nutriments minéraux. Il comprend le métaxylème, qui se développe après le protoxylème.
- Fonction : Le xylème mature transporte l’eau et les sels minéraux des racines vers les feuilles et contribue également au soutien mécanique de la plante grâce à ses parois cellulaires épaisses et lignifiées.
- Caractéristiques : Le xylème est composé de plusieurs types de cellules, y compris des trachéides, des éléments de vaisseau, des fibres xylémiques et des cellules parenchymateuses, offrant à la fois le transport et le soutien structurel.
Protophloème
- Définition : Le protophloème est la première phase de développement du phloème, le tissu vasculaire chargé du transport des sucres et d’autres molécules organiques dans la plante.
- Fonction : Situé dans les zones de croissance de la plante, le protophloème transporte les nutriments des zones de photosynthèse vers les parties en croissance. Ses cellules peuvent être plus petites et moins spécialisées que celles du phloème mature.
- Caractéristiques : Les éléments du protophloème sont souvent les premiers à devenir fonctionnels mais peuvent être écrasés ou devenir obsolescents à mesure que la plante mûrit et que le phloème mature se développe.
Phloème
- Définition : Le phloème est un tissu vasculaire mature qui transporte les produits de la photosynthèse (sucres) et d’autres substances organiques à travers la plante.
- Fonction : Il joue un rôle crucial dans la distribution des nutriments produits dans les feuilles vers toutes les parties de la plante, y compris les racines.
- Caractéristiques : Le phloème mature comprend plusieurs types de cellules, notamment les tubes criblés, les cellules compagnes, les fibres phloémiennes et le parenchyme phloémien, chacun jouant un rôle spécifique dans le transport et le stockage des nutriments.
En résumé, le protoxylème et le protophloème représentent les étapes initiales de développement du xylème et du phloème, essentiels pour le transport de l’eau, des minéraux et des nutriments dans les plantes, tandis que le xylème et le phloème matures sont des versions plus avancées et structurées de ces tissus.